


提出生物布林根(Bouligand)结构形成的全新自组装机制
近日,华中农业大学工学院刘培文研究员研究成果在Nano Today发表。研究在设计的甲壳素纳米晶在限域空间中的挥发自组装中发现了全新的固定界面挥发自组装模式(FB-EISA),该模式下所形成微观结构中的变化取向梯度结构以及自组装过程对复杂环境的强稳定性,极好的扫清了目前挥发自组装策略在被用来解释生物体三维功能结构形成过程中的阻碍,特别为理解Bouligand结构的天然形成过程进而重新设计与构建类似的精细结构提供了新思路。
甲壳素的天然纳米单元在三维空间上有序排布所形成的Bouligand结构广泛存在于天然生命体内,为许多生物的坚固力学以及多彩的生物光学奠定了结构基础。研究者们一直希望能够利用合适的机制去解释Bouligand结构的天然形成过程进而重新设计与构建类似的精细结构,其中,自组装策略被认为极具潜力。然而,目前利用自组装过程来合理地解释生物体内Bouligand结构的形成依然是一个极大的挑战。
以往研究发现,利用传统的挥发诱导甲壳素纳米晶(或其它类似的一维纳米材料-纤维素纳米晶)自组装来获得类似的结构时,常存在以下三个问题,即所得到的结构常含有大量的缺陷,这导致其光学结构常存在混乱的颜色分布;所得结构大尺度上结构单一(平行结构),与自然界中的带有取向变化的梯度结构不相符合;现有的自组装过程对环境稳定性的要求苛刻,这与复杂自然条件下的生命体功能结构的构建不吻合。其主要原因是目前所使用的自组装模式存在固有的缺陷。传统的挥发诱导自组装(Classical EISA),其挥发面与固体结构的初始形成面是分离并且不稳定的(图1)。这导致在挥发的过程中,挥发面的溶质或者纳米颗粒会因为挥发而形成大量分散的、具有有序堆叠结构的小颗粒(Tactoids)。这些小颗粒因为密度高于周边溶液,会逐渐沉淀到底部并堆积融合,造成缺陷结构。因此,为了得到足够均匀的结构,在传统的挥发诱导自组装的过程中,一般会要求足够长的自组装时间以及基本不变的自组装环境。
 图1-传统的挥发诱导自组装模式VS该工作提出的新的挥发自组装机制
图1-传统的挥发诱导自组装模式VS该工作提出的新的挥发自组装机制研究工作人员受到昆虫甲壳颜色结构是由一个个极小的颜色单元构成的事实启发,创造性的将甲壳素纳米晶的挥发自组装在一个极小的空间-规则毛细管(圆形或者正多边形)内进行,从而甲壳素纳米晶悬浊液在挥发自组装的过程中表现出了与传统挥发诱导自组装完全不一样的特征,即甲壳素纳米晶悬浊液的挥发面锚定在毛细管的末端,另一个在毛细管内部的甲壳素纳米晶悬浊液液面不断的地向锚定在毛细管末端的挥发面移动,这使得挥发面与初始沉积面实现了重合,并且两者一直保持着稳定的状态(图2A)。

 图2-甲壳素纳米晶限域自组装的双折射结构
图2-甲壳素纳米晶限域自组装的双折射结构在限域挥发诱导自组装的过程中,甲壳素纳米晶悬浊液以及所形成的有序结构都被使用偏振光显微镜观测,发现甲壳素纳米晶因为挥发被诱导形成的有序结构在偏振光显微镜下显示为梯度、多彩以及动态的两种双折射结构:一种是多层抛物面结构,另外一种是嵌套多层的管状双折射结构(图2)。经过观测,挥发面与初始沉积面的双折射结构在自组装的全过程中一直显示为动态状态,表现为动态颜色变化。在横截面与纵截面的双折射结构的颜色过渡部分有明显的颜色界限,没有发现杂乱排布的双折射颜色,这意味着所形成的微观结构是高度规则的。另外,在形成高度有序结构区域与甲壳素纳米晶非有序结构区域的分界处未发现甲壳素纳米结构的聚集体(Tactoids),这表明报道的限域挥发诱导自组装与前面发现的传统的挥发诱导自组装过程不同。
 图3-甲壳素纳米晶限域自组装所形成的微观结构
图3-甲壳素纳米晶限域自组装所形成的微观结构同时,甲壳素纳米晶经限域挥发诱导自组装所形成的微观结构也被Micro-CT与SEM进行了详尽的观测。与传统挥发诱导自组装中只有平行的层状结构不同,本工作所得的微观结构是一个三维的梯度密度结构,从挥发面存在的毛细管末端向内,层状的甲壳素纳米晶结构的密度越来越小。另外,在同一个横截面上,层状结构的密度也随着与中心的距离变短而变小。同时,在中间态的固体结构中,发现本自组装的层状结构不仅仅只是平行结构,而是有多维度的变化,这与自然界中昆虫有一定的变化层状结构相符合。
 图4-该限域自组装对复杂环境优异的稳定性
图4-该限域自组装对复杂环境优异的稳定性传统的自组装过程都对环境的稳定性有着严苛的要求,哪怕细微的干扰都将对自组装过程有不可控制的干扰,导致最后得到的结构有极大的变化。该工作中的限域挥发诱导自组装(FB-EISA)对温度的提高、震荡以及与重力的夹角变化都不敏感(图4)。干扰状态FB-EISA所形成的结构,与无干扰状态相比,基本保持不变。表现出在复杂环境下优异的稳定性,为生物体Bouligand结构的构建提供一种可信的解释途径。经过作者仔细的研究以及推导,该FB-EISA行为如此特殊特征的原因如下(图1B):挥发面固定在毛细管的末端,该挥发面同时作为初始沉积面;在整个系统中,相界面只有一个,完整并且连续,并且不是平面的,水的挥发需要从低浓度区向高浓度区渗透,通过高度有序结构后挥发出去,这使得存在的缺陷结构有足够的机会去调整达到能量最低的平衡状态;甲壳素纳米晶的生长不是Tactoids堆积,而是在相界面上的局部脱水相变,前面所形成的有序结构称为后续有序结构的生长提供了模版。
该全新的挥发诱导自组装模式的发现对理解天然Bouligand结构的形成、构建均匀的纳米功能结构以及获得较大的有机、高分子与蛋白质晶体具有重要的意义。
华中农业大学为第一单位,刘培文研究员为论文第一作者,安徽大学王加秀教授,东北林业大学戚后娟博士为共同第一作者,哥廷根大学Kai Zhang教授为通讯作者。
论文链接:https://doi.org/10.1016/j.nantod.2022.101420

揭示水稻垩白形成的分子机理
近日,华中农业大学作物遗传改良国家重点实验室何予卿教授团队的研究论文在The Plant Cell杂志在线发表,揭示了水稻心白率基因WCR1的自然变异和胚乳氧化还原稳态影响垩白形成的分子机理。
垩白(Chalkiness)是由于稻米胚乳中淀粉颗粒和蛋白体排列不紧密而产生的白色不透明的部分,全面影响稻米外观和蒸煮食味品质,直接影响稻米作为商品的市场价值。因此,降低垩白是水稻优质遗传改良的全球共同育种目标。垩白是典型的数量性状,受遗传因子和环境因素的共同影响,这也使得垩白基因克隆面临了很大的挑战。目前,通过自然群体克隆的垩白相关基因非常少,垩白形成的分子机制也有待进一步探索。

A: WCR1的图位克隆;B:近等基因系株型和籽粒垩白表型;C:WCR1参与垩白形成的调控模式。
该研究经过多年的图位克隆,在水稻第一染色体上定位到一个控制心白率的基因WCR1,WCR1BL位点除降低心白率外,同时还增加产量和米饭食味值。系统发育和序列分析表明该基因编码一个F-box蛋白,在大麦、小麦、玉米、高粱和拟南芥中都存在同源基因。WCR1是心白率的负调控因子,上调WCR1的表达水平,能显著降低心白率。自然变异分析显示,启动子上预测的一个功能性变异位点A/G可能是引起表达差异和心白率差异的重要原因。通过系统发育分析,表明WCR1G等位型来源于野生稻,而WCR1A则主要存在于热带粳稻中。进一步研究发现,A/G变异引起了OsDOF转录因子结合位点核心序列的改变,通过酵母单杂、瞬时表达和EMSA实验验证了OsDOF17与WCR1启动子存在互作,并且A/G变异会影响OsDOF17的结合能力。
为了解析WCR1调控垩白的分子机制,通过酵母筛库、BiFC以及Pull-down等实验筛选到一个WCR1的互作蛋白MT2b。MT2b编码一个金属硫蛋白,具有结合金属离子和清除活性氧(ROS)的作用。研究者从遗传、生理和生化上证明了WCR1具有调控MT2b转录水平和抑制26S蛋白酶体介导MT2b蛋白降解的功能,从而促进ROS清除和延迟胚乳细胞程序性死亡(PCD),最终增加了贮藏物质的积累和降低心白率。该项工作不仅阐释了心白形成的分子机制,同时还解析了垩白的自然变异,挖掘了优势等位基因,为稻米品质的改良提供了理论基础和具有应用前景的基因资源。
WCR1是该团队继Chalk5(Li et al, Nature Genetics, 2014)克隆后的又一重要发现,华中农业大学作物遗传改良国家重点实验室博士生吴边为本文第一作者,何予卿教授为本文的通讯作者,姚家玲教授和周竹青教授参与了本研究部分工作。
论文链接:
https://doi.org/10.1093/plcell/koac057
构建首个棉花纤维高分辨率动态3D基因组结构图谱
近日,华中农业大学棉花遗传改良团队首次构建了棉花纤维的高分辨率三维基因组结构图谱,揭示了亚基因组协作调控异源四倍体棉花纤维发育的拓扑结构基础,对棉花功能基因组研究具有重要推动作用。同时,棉花纤维作为纯净的植物单细胞类型,该研究为解析其他植物单细胞分化的转录调控机制提供了参考。相关研究成果发表在国际学术期刊Genome Biology上。
植物细胞分化伴随着基因转录调控网络重塑,该过程受到表观遗传修饰和染色质结构等影响。棉花纤维是胚珠表皮上高度分化的单细胞结构,其分化发育过程一般经历约50天,分为四个相对重叠阶段:起始分化、伸长、次生细胞壁合成和脱水成熟期。因此,棉花纤维可作为研究植物细胞分化的模式系统。该团队前期的研究发现,随着纤维发育,CHH (H = A、T 或 C) DNA 甲基化水平逐渐增加,伴随着常染色质向异染色质的转换(Nucleic Acids Research, 2016)。随着三维(3D)基因组学技术的发展,该团队利用棉花叶片进行比较三维基因组研究,发现了基因组多倍化和基因组大小演化中染色质高级结构的重组规律(Nature Plants, 2018; Molecular Biology and Evolution, 2021)。
受制于棉花纤维细胞的生理特殊性,普通的Hi-C等实验效果不理想,纤维的三维基因组研究进展缓慢。本研究通过对原位Hi-C技术进行改进,极大地提高了Hi-C实验数据的信噪比。研究中利用海岛棉3-79开花后0天(DPA)、5天(DPA)、10天(DPA)、20天(DPA)的胚珠或纤维样品——分别代表纤维起始分化、伸长(早期伸长和快速伸长)和次生壁合成阶段,进行纤维的三维基因组学研究,旨在解析纤维发育中的染色质拓扑结构改变与基因转录调控之间的关系。
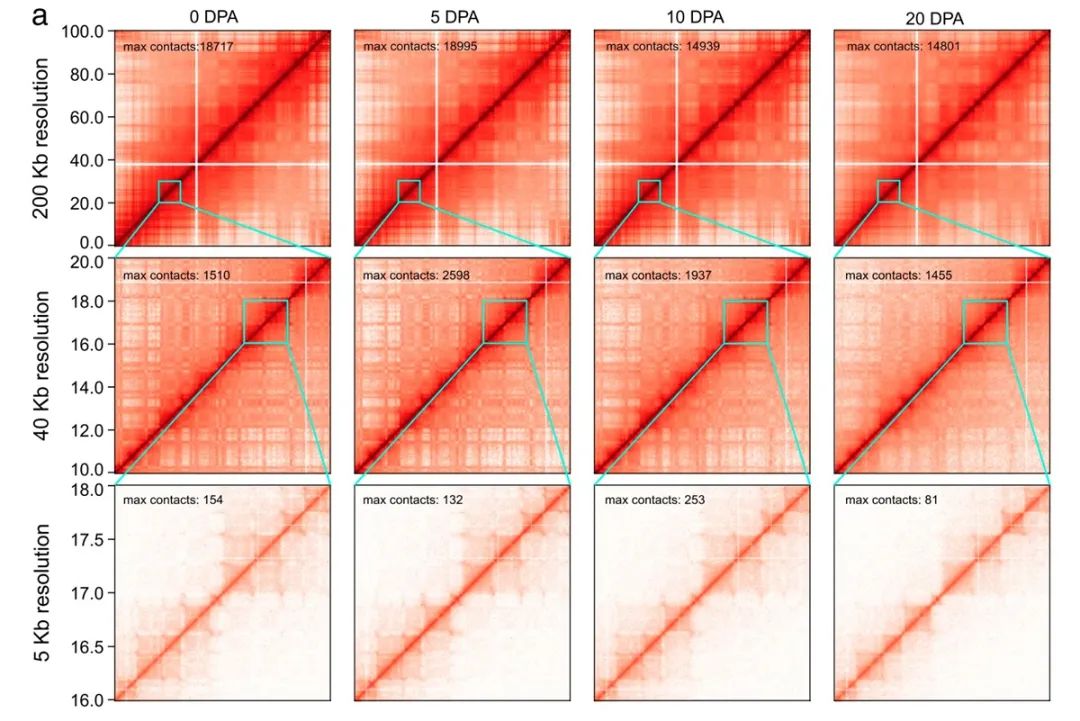 图1. 不同分辨率下棉花纤维的三维基因组结构图谱
图1. 不同分辨率下棉花纤维的三维基因组结构图谱研究发现,随着纤维发育,活跃表达的基因变少,同时活性染色质修饰减少,并且At和Dt两个亚基因组对异源四倍体纤维的发育存在协同贡献。在纤维发育过程中,不同的分辨率下纤维细胞核内呈现了不同的染色质高级结构(图1);染色质区室从活跃到不活跃的转换伴随着纤维发育相关基因的沉默。研究鉴定了10571个拓扑关联结构域 (TAD),其中25.6%在不同发育阶段中特异存在,75.23%在两个亚基因组之间发生分割或融合,而这种亚基因组间同源TAD的结构变化与转座子的插入/消除有关。值得注意的是,部分远程 TAD-TAD 相互作用组成的复杂 TAD 团(clique)在纤维发育后期逐渐解散,这是后期染色质高级结构变化的一个突出特征。染色质环介导的转录调控网络在纤维发育过程中发生显著变化,而且在两个亚基因组之间也存在明显差异,这可能与同源基因的偏向性表达相关(图2)。这项研究揭示了四倍体棉花纤维发育中两个亚基因组的时空不对称染色质结构,并为植物细胞阶段性分化的调控机制提供了新的见解。
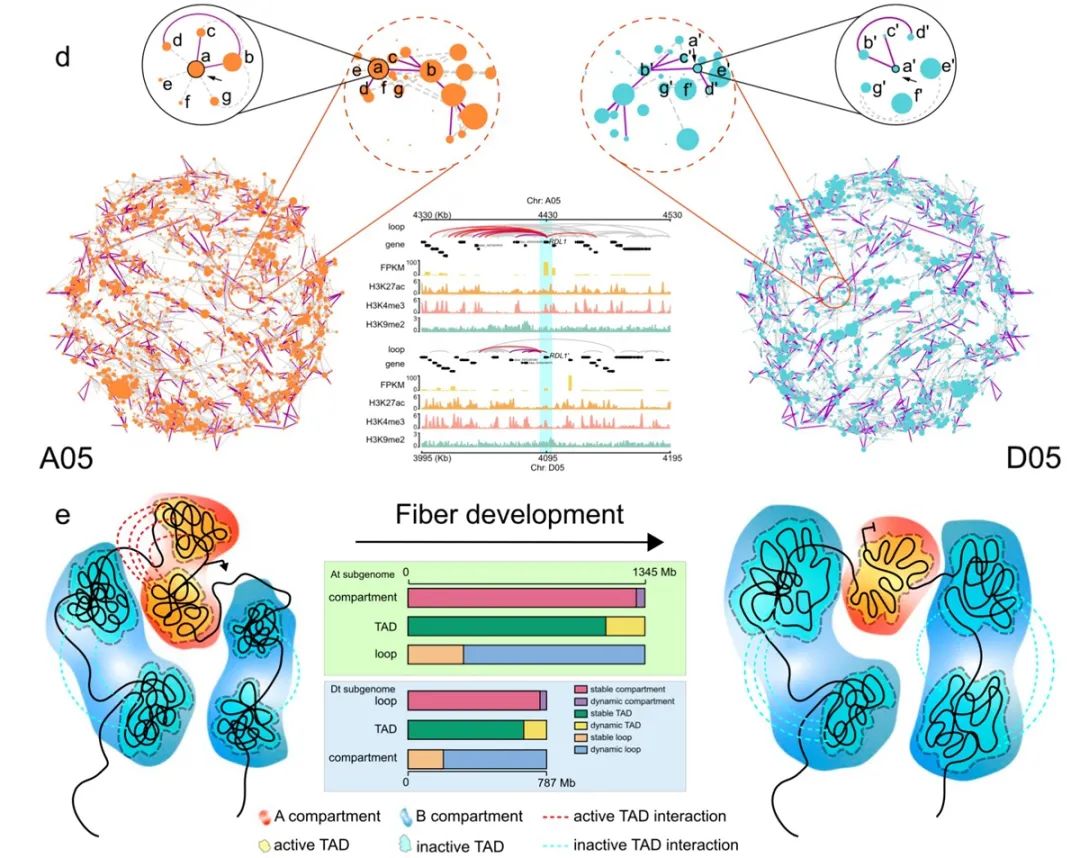 图2. 染色质环介导At和Dt亚组间差异性的调控网络
图2. 染色质环介导At和Dt亚组间差异性的调控网络作物遗传改良国家重点实验室、湖北洪山实验室博士生裴柳玲和黄鲜晖为论文共同第一作者,王茂军教授为通讯作者,张献龙教授参与了课题设计。
论文链接:
https://doi.org/10.1186/s13059-022-02616-y
版权声明
本文仅代表作者观点,不代表本站立场。
本文系作者授权发表,未经许可,不得转载。
本文地址:http://www.shanxidayuan.com/zcjh/jishixinwen/52547.html









